 5 X5 \! X& e- Q& u! E5 M. f
5 X5 \! X& e- Q& u! E5 M. f 点击上方 环境微生物组学 关注我们  , T2 q' P; {9 F+ W. B7 @: u. W
, T2 q' P; {9 F+ W. B7 @: u. W -研究背景-
; G6 t$ z9 Z/ q, R( u8 ? 地球的 70% 被海洋覆盖,海洋地壳是最大的地下环境,表明它有潜力成为微生物生命的大储藏库。由于海洋地壳相对难以接近,几乎没有机会了解地壳微生物的生命及其代谢过程。
$ a/ C; O. n# g$ `: d% c& T 在Juan de Fuca Ridge (JdFR)的东侧发现了一个经过充分研究的海底洋壳环境,由3.5Ma玄武岩组成。对海底钻孔观测站进行的三十年研究揭示了海底流体流动和流体-岩石-微生物相互作用的动态特性。然而,对于特定微生物群如何在这种还原剂耗尽的栖息地中生存,仍存在不确定性。在这种环境中,一个经常观察到但神秘的群体是Acidobacteriota的Aminicenantia纲,以前称为Obsidian Pool 8 (OP8 )。此类已在多种缺氧环境中发现,但不存在培养代表。Aminicenantia调查在缺氧水生环境中,证明了复杂碳水化合物和蛋白质的降解和发酵、Wood-Ljungdahl 驱动的乙酸盐氧化或通过甲酸盐氧化产生CO2的基因组潜力。这些研究得出结论, Aminicenantia的生态作用可能是清除和分解有机物。与上层海水 (39μM DOC) 相比,JdFR 侧翼的缺氧地壳流体含有低浓度的有机碳 (12μM DOC),这引发了关于哪种代谢策略Aminicenantia的问题习惯在这种环境下生活。
# P z& s( w) Q k5 X; [( w 先前的工作基于宏基因组解释,在 Juan de Fuca Ridge 侧翼的地下海洋地壳中确定了两个不同的Aminicenantia进化枝。在这里,这项先前的工作被用作基础,通过使用公开可用的和新的宏基因组和单细胞基因组数据集的组合来研究在海洋地壳中持续存在的神秘未开垦的Aminicenantia的潜在代谢特征。我们的数据揭示了 19 种JdFR Aminicenantia的生活策略,它们形成了两个谱系,JdFR-78 和 JdFR-80,在这个低能量栖息地中,提供了海底是早期进化谱系储存库的证据,并暗示了可能的培养方法。 4 E: b- \! ]& C; l+ @' n$ Z

, D3 l0 a+ M8 |- n5 u. ^( M, ` -研究结果- - @7 g. n* s P$ Z5 T Z; l/ H
研究结果发现,两种谱系均通过多种机制产生 ATP,包括底物水平磷酸化、厌氧呼吸和驱动 Rnf 离子易位膜复合物的电子分叉。基因组比较表明,这些Aminicenantia将电子转移到细胞外,可能转移到与该地点的矿物学一致的铁或硫氧化物。 JdFR-78 的谱系具有小型基因组,这些基因组是 Aminicenantia 的基础类并可能使用“原始”siroheme 生物合成中间体进行血红素合成,表明该谱系保留了早期进化生命的特征。谱系 JdFR-78 包含 CRISPR-Cas 防御系统,而其他谱系包含可能有助于防止超级感染或无法检测到病毒防御的前噬菌体。总的来说,基因组证据表明Aminicenantia通过利用简单的有机分子和细胞外电子传输很好地适应了海洋地壳环境。
+ B+ H2 N+ M# n: @' Z( @0 ~" { 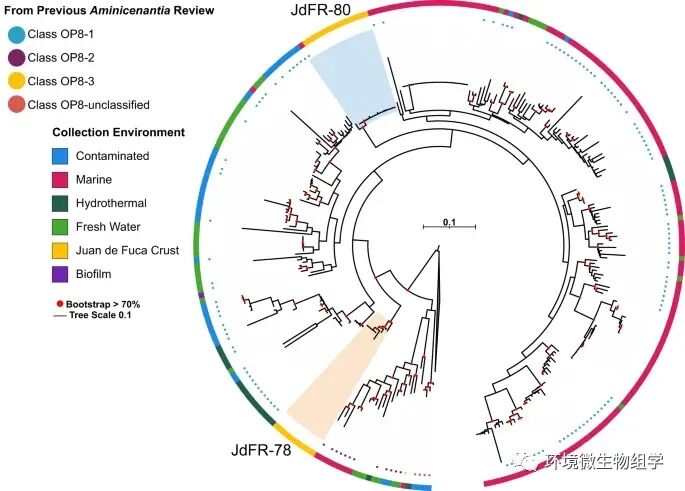
% R% B7 r' k0 {+ ]# x% i# D3 U. A$ O
图1. Aminicenantia 16S rRNA 基因系统发育树揭示了生活在 Juan de Fuca Ridge 侧面洋壳中的两个谱系 9 F; O; Y" G" `/ T/ L3 r; z
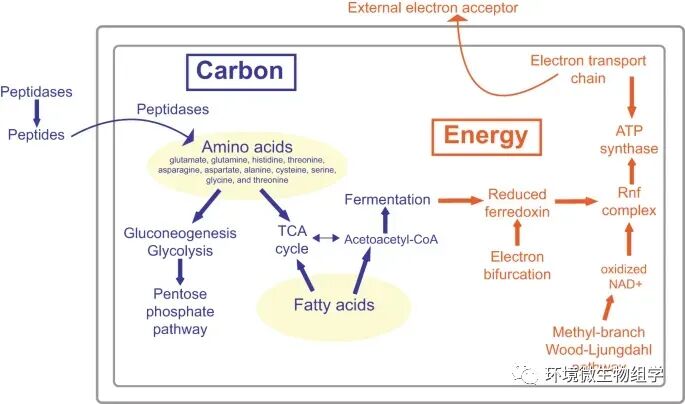
- _9 D5 [ f6 L3 f# C4 V
图 2:Juan de Fuca Aminicenantia中的碳利用和能量产生代谢
% p( K8 _! K- u! f0 H. D 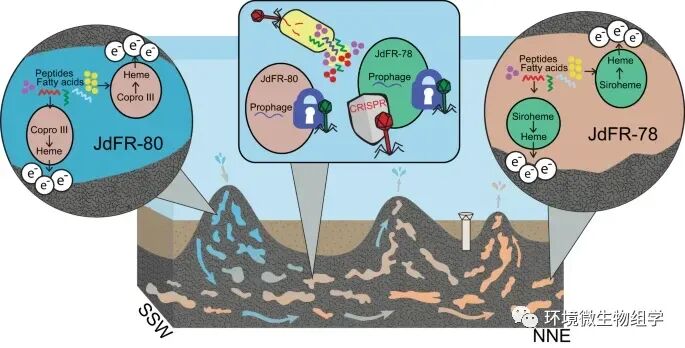
8 T3 M1 }/ G$ i. _+ a; ?! w9 u
图 3:Aminicenantia的两个谱系的生活策略 / T* e* S/ I1 [" Z8 k: |
更多图形图例、研究内容,推荐阅读原文 (https://doi.org/10.1038/s41467-023-38523-4) ! - m" ~& w: h8 e* i
如若侵犯到原作者任何相关利益,请告知删除!翻译过程存在不准确或表述不清,以及任何疏漏,欢迎大家后台/邮件(Env_multiomics@outlook.com)留言指正。
; v7 { G- b t" L; q 点个在看你最好看  6 ~, g2 z! w8 A1 N& q2 Z+ Q; p0 j2 l: P( s
- m5 ?' C% X8 _
( C" s( ?' ?8 j; N# G% \( e- M- R. y' E, O4 j% j0 `5 T
6 ~, g2 z! w8 A1 N& q2 Z+ Q; p0 j2 l: P( s
- m5 ?' C% X8 _
( C" s( ?' ?8 j; N# G% \( e- M- R. y' E, O4 j% j0 `5 T
| 

